| |
La
extraordinaria capacidad osmorreguladora
Algunas
especies de anostracos son eurihalinas y pueden vivir
en aguas salobres o hasta saladas e hipersalinas. En
los casos más extremos, las especies de Artemia y Parartemia toleran
asombrosas variaciones en la salinidad del medio de
20-30% hasta 300-330% (Grassé, 1996) y presentan
un comportamiento osmorregulador complejo. En ambientes
salinos diluidos, cuando su biotopo está inundado, Artemia (Fig.
2) hace una regulación hiperosmótica,
bombeando iones hacia su medio interno y excretando
una abundante orina muy diluida, para elevar la osmolaridad
de su hemolinfa por encima de la del medio; en agua
con una concentración cercana a la del agua
de mar, se comporta como un osmoconformador, manteniendo
sus fluidos corporales con una osmolaridad similar
a la del medio; finalmente, en salmueras concentradas,
cuando las lagunas saladas se están evaporando,
osmorregula hipotónicamente, bombeando iones
desde la hemolinfa hacia el medio externo y excretando
una orina muy concentrada (hasta 4 veces por encima
de la concentración de la hemolinfa) (Barnes,
1977; Purves y col., 2004). Sin embargo, probablemente
asociado con su notable indefensión a la predación,
no existen anostracos marinos.
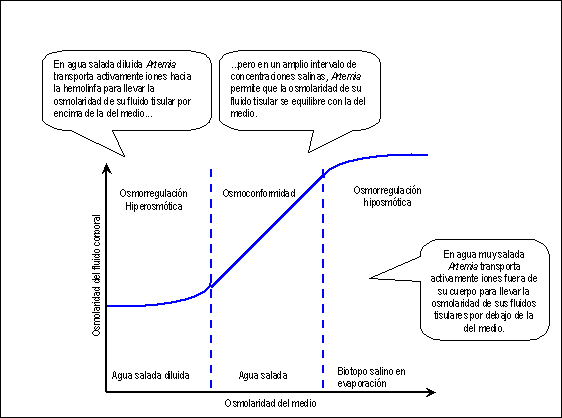
Figura 2. Los biotopos pueden variar mucho
en concentración salina. Artemia presenta
una gran capacidad adaptativa: actúa como
regulador hiperosmótico en aguas saladas diluidas,
como osmoconformador en concentraciones salinas medias
y como regulador hipoosmótico en aguas muy
saladas. (Adaptado de Fig. 51.1 Purves y col., 2004)
Las
estructuras osmorreguladoras
Las larvas de los anostracos realizan la regulación
osmótica de la hemolinfa, intercambiando iones
con el medio a través del órgano nucal
(Hootman y Conte, 1975). Esta estructura, mencionada
en la bibliografía como ‘neck organ’,
‘nuchalorgan’, ‘dorsal organ’,
‘neck shield’, ‘salt gland’,
‘calotte cervicale’, ‘tubercule post-oculaire’,
se ubica dorsalmente, por detrás de los ojos,
en la región cefálica de las larvas de
los anostracos; también está
presente en larvas de los otros branquiópodos
así como en las de Rehbachiella kinekullensis Müller1.
Según Waloßek (1995) el órgano
nucal se habría originado a partir de un órgano
sensorial primordial en la misma posición, distal
al tercer segmento cefálico con apéndices.
Este órgano persiste al menos hasta cierto punto
del desarrollo larval y puede degenerar o permanecer,
probablemente no funcional, hasta antes de alcanzar
la adultez (Martin, 1992). Las larvas tendrían
una mayor capacidad para transportar iones que los
adultos, por lo que tolerarían mayores intervalos
de salinidad.
Durante el desarrollo post-embrionario el órgano
nucal es progresivamente reemplazado en su función
osmorreguladora por los epipoditos de los toracópodos,
y en menor grado por la pared intestinal (Croghan,
1958; Grassé, 1996). En el adulto, el epitelio
de los epipoditos es la principal sede de intercambio
iónico contra gradiente: allí se incorporan
activamente iones del medio a la hemolinfa en la regulación
hiperosmótica o se los elimina activamente desde
la hemolinfa hacia el medio externo en la regulación
hiposmótica.
|
|
