Vol. 24, núm. 1 enero-febrero 2023
Escarabajos vagabundos: nuestros aliados invisibles en el suelo
Diana María Méndez Rojas, Carlos Andrés Cultid Medina y Federico Escobar CitaResumen
Muchos escarabajos que habitan el suelo pasan desapercibidos al ojo humano debido a su pequeño tamaño y colores poco llamativos. Este es el caso de los escarabajos conocidos como vagabundos, de la familia Staphylinidae. Dichos escarabajos son comunes en muchos ecosistemas terrestres y se les encuentra asociados principalmente a la hojarasca; también viven bajo piedras y troncos, o son errantes sobre el suelo. Este grupo de escarabajos desempeña un papel funcional como depredadores de otros invertebrados, tanto en ecosistemas naturales, como en los sistemas agrícolas. En el escrito, destacamos su enorme diversidad, las múltiples interacciones ecológicas que establecen con otras especies y su importancia para la salud de los ecosistemas.
Palabras clave: insectos, depredadores, macrofauna del suelo, estafilínidos.
Rove beetles: our invisible allies on the ground
Abstract
Many ground-dwelling beetles are unnoticed by the human eye due to their small size and inconspicuous colors. This is the case of rove beetles that belong to the Staphylinidae family. These beetles are common in many ecosystems and are associated with leaf litter, live under rocks and logs, or are walking on the ground. In the work we show how this group of beetles plays a functional role as predators of other invertebrates both in natural areas and in agricultural systems. In addition, we highlight its enormous diversity, the multiple ecological interactions established with other species and its importance for the health of ecosystems.
Keywords: insects, predators, ground macrofauna, staphylinids.
¿Qué son los estafilínidos y cómo es posible reconocerlos?
Los escarabajos conocidos como “vagabundos” o “errantes” son coleópteros que pertenecen a Staphylinidae, una de las familias de insectos más diversas a nivel mundial con cerca de 63 mil especies descritas (Irmler y Lipkow, 2018). La mayoría de los estafilínidos se diferencian fácilmente de escarabajos de otras familias por presentar las antenas filiformes (i.e., antenas en forma lineal sin presentar mazo), y el primer par de alas cortas y endurecidas (conocidas también como élitros), dejando el abdomen expuesto y flexible (Parker, 2017, figura 1a). Aunque podemos encontrar excepciones, algunas especies de las subfamilias Omaliinae, Scaphiidinae y Scydmaeninae presentan los élitros alargados cubriendo la mayor parte del abdomen. En general, los estafilínidos pueden mover su abdomen y plegarlo o recurvarlo hacia arriba de forma similar a como lo hacen los alacranes o los insectos conocidos como tijerillas (orden Dermaptera, figura 1b), con los cuales son comúnmente confundidos (Navarrete-Heredia et al., 2002).

Figura 1. Platydracus fulvomaculatus, una especie de escarabajo estafilínido (a, © Ricardo Arredondo) donde muestra como estos escarabajos pueden plegar el abdomen y ser comúnmente confundidos con las tijerillas (b, orden Dermaptera, © Paul).
Fotografías tomadas de www.naturalista.mx
Si bien la gran mayoría de los representantes de estos escarabajos tienen el cuerpo plano y delgado (ver figura 2, panel 2.1, 2.9), ellos exhiben una gran diversidad de formas: algunas especies tienen el abdomen cilíndrico (figura 2, panel 2.2, 2.5), mientras que otros son de cuerpo redondo y compacto (figura 2, panel 2.4, 2.7). Contrario a lo observado en otros grupos de escarabajos, el común de los machos de los estafilínidos no exhiben estructuras morfológicas llamativas, como grandes cuernos, mandíbulas enormes o patas extremadamente largas que los diferencien de las hembras. En algunas especies la diferenciación sexual de estafilínidos es muy sutil, se observa principalmente en machos con un tamaño de la cabeza mayor y mandíbulas un poco más grandes que las hembras, en modificaciones de las patas traseras e incluso en la presencia de pequeños cuernos en la cabeza (Navarrete-Heredia et al., 2002). Descrito lo anterior, el objetivo de este artículo es dar a conocer la existencia del grupo de seres vivos más numeroso de nuestro planeta: los estafilínidos.
¿Por qué no los vemos?
A diferencia de otros insectos vistosos y comunes en los bosques, cultivos y zonas urbanas, los estafilínidos son en general de tamaño pequeño (6 mm), de coloración oscura y opaca, principalmente de color marrón o negro, lo que dificulta su observación y por esta razón pasan desapercibidos ante la vista humana. Los estafilínidos, junto con las arañas y las hormigas, dominan en los suelos de los bosques y los campos agrícolas, donde las especies suelen vivir en la hojarasca y en los troncos en descomposición, sitios donde predominan alta humedad y bajas temperaturas (figura 2). Además, el común de las especies son de hábito nocturno, lo que significa que se esconden durante el día y salen en búsqueda de alimento o pareja cuando se oculta el sol (Parker, 2017; Frank y Thomas, 2002).

Figura 2. Infografía donde se ilustra la variación de formas y tamaños que tienen los estafilínidos. El suelo, la hojarasca, la vegetación, las orillas de ríos y arroyos son los hábitats más comunes en los que viven los estafilínidos.
¿Por qué son tan exitosos?
Los estafilínidos tienen innumerables estrategias de vida que les ha permitido colonizar diversos ambientes, desde el nivel del mar hasta la cima de las montañas, en alturas superiores a 4000 m. Los biólogos sugieren que el gran número de especies de escarabajos errantes se debe, en parte, a que presentan altas capacidades de movimiento ya que son buenos voladores y caminadores, lo que les ha permitido conquistar y explorar distintos ambientes (Pohl et al., 2007). Aunque la mayoría de las especies dominan el suelo y están asociadas a cadáveres de animales y a restos vegetales, muchas otras viven en nidos de hormigas, termitas, aves y mamíferos. También se conocen especies que viven asociados a hongos o en lugares muy húmedos cerca de ríos y arroyos (Frank y Thomas, 2002). Debajo de la hojarasca, directamente en el suelo, también es posible encontrar a los estafilínidos menos conocidos y de menor tamaño (1.8 mm), que al parecer se alimentan de organismos microscópicos (Navarrete-Heredia et al., 2002).
El grueso de las especies de estafilínidos son depredadoras voraces de otras especies de invertebrados, algunas más se alimentan de hongos y son conocidas como micófagas, otras son fitófagas, se alimentan de tejido vegetal vivo, y las hay saprófagas, es decir que se alimentan de materia vegetal o animal en descomposición. Existen especies de estafilínidos cuyas larvas se alimentan y desarrollan en el interior de otras larvas o pupas de moscas, esta estrategia se conoce como parasitoidismo, como es caso del género Aleochara (Navarrete-Heredia et al., 2002).
¿Son nocivos para los seres humanos?
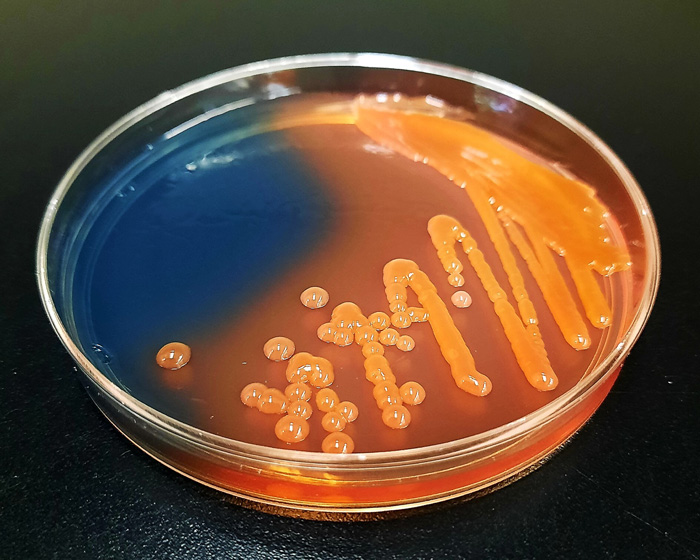
Casi el total de los estafilínidos son completamente inofensivos, aunque hay pocas especies conocidas que pueden llegar a ser nocivas para los seres humanos, entre ellas se encuentran algunas especies conocidas como bichos de fuego, culebrillas, panchos o picabueyes, pertenecientes al género Paederus (figura 3). Estos pequeños escarabajos habitan en todo el mundo y suelen tener una coloración naranja con los élitros azules. Si se les molesta o se les aplasta contra la piel humana, esos escarabajos segregan una sustancia irritante que causa una aguda dermatitis de contacto (Silva et al., 2014). La sustancia es conocida como pederina, una potente toxina que produce ampollas y es usada como mecanismo de defensa contra sus enemigos naturales y se encuentra presente tanto en los huevos, como en las larvas y en los adultos; cuando dicha sustancia entra en contacto con la piel humana se produce una lesión en forma de vejiga, que viene acompañada con una sensación de ardor, dolor y escozor (Silva et al., 2014). En México se conocen diecinueve especies de culebrillas, pero hasta ahora sólo se ha reportado un único caso de dermatitis en el estado de Jalisco (Navarrete-Heredia et al., 2002).

Figura 3. Escarabajos estafilínidos pertenecientes al género Paederus conocidos como panchos o picabueyes (Izquierda, © Felix Riegel; derecha © Gerónimo Martín Alonso).
Fotografías tomadas de www.naturalista.mx
Escarabajos errantes, hormigas y termitas: una relación huésped y anfitrión
Varias especies de estafilínidos son habitantes estrictos en nidos de hormigas (figura 2, panel 4), mientras que otras usan los desechos de los nidos como fuente de alimento (Márquez y Navarrete-Heredia, 1994; Navarrete-Heredia et al., 2002). A las especies que viven en las colonias de otras especies se les denomina parásitos sociales. A través de cambios dramáticos de la forma del cuerpo, el comportamiento y la imitación química de sustancias de comunicación conocidas como feromonas, los estafilínidos parásitos ganan la aceptación de las hormigas y se aprovechan de ellas. Así, los escarabajos pasan como impostores, ya que se parecen, caminan, huelen y se comportan similar a las hormigas. Una vez que los escarabajos son aceptados en las colonias, roban en secreto la comida de las hormigas y se comen sus huevos, engañándolas para alimentarse con las crías de la colonia. Incluso algunas especies de estafilínidos se reproducen dentro de los nidos y hacen que las hormigas alimenten y críen a sus propias larvas (von Beeren et al., 2018). Estrategias similares han sido observadas en los estafilínidos que viven en los nidos de termitas en distintas regiones del mundo. Esta asociación es conocida como termitófila, ya que las especies están morfológicamente especializadas para vivir en los nidos de las termitas y pasar al menos un estado completo de su ciclo de vida (Moreira et al., 2019).
Pequeños pero importantes
Dada su alta diversidad y sus variadas formas de vida, los escarabajos son elementos claves de las redes tróficas en cultivos y pastos. La importancia ecológica de estos escarabajos en los ecosistemas es incalculable y hasta ahora poco estudiada, especialmente en las regiones tropicales.
Los estafilínidos pueden llegar a ser cinco veces más abundantes que otros artrópodos depredadores, como es el caso de las arañas (Klimaszewski et al., 2018), por lo que también son considerados depredadores importantes en los ecosistemas. Sus presas suelen ser otros insectos e invertebrados que habitan en el suelo (Good y Giller, 1991). En general, los escarabajos de menor tamaño suelen consumir huevos de otros insectos, mientras que escarabajos más grandes depredan a las larvas y los adultos (Halimov, 2020). Por su parte, el común de los estafilínidos depredadores que viven en el suelo y están asociados a la hojarasca (figura 4) son considerados como potenciales agentes de control biológico en cultivos de cereales, como el trigo, la cebada y el maíz (Bohac, 1999; Klimaszewski et al., 2018).

Figura 4. Estafilínido depredador localizado frecuentemente en el suelo (subfamilia Staphylininae).
Foto © David Hoyos Velásquez.
Algunas, aunque pocas especies de escarabajos, también se alimentan de partes florales, polen y néctar; otras son polinizadoras, en particular de un conjunto de plantas como los anones, las magnolias y las aráceas (Sayers et al., 2019). Asimismo, ciertos estafilínidos forman congregaciones en las brácteas florales de plantas conocidas como heliconias (Heliconiaceae) y columneas (Gesneriacea), donde depredan larvas y pupas de moscas o se alimentan de materia orgánica en descomposición (López-García y Marín-Gómez, 2018). Además de ser en su mayoría escarabajos depredadores, hay un grupo que se alimenta de materia vegetal o animal en descomposición, es decir, son saprófagas, encontradas comúnmente en la hojarasca, en troncos en descomposición, sobre hongos o utilizando recursos efímeros como las brácteas florales, frutos en descomposición o cadáveres de animales (Navarrete-Heredia et al., 2002; Parker, 2017). Lo anterior implica que diversos estafilínidos contribuyen al reciclado y redistribución de nutrientes en el suelo.
¿Qué acciones humanas pueden afectar a estos escarabajos?
Las actividades humanas productivas han transformado el planeta y han cambiado la dinámica ecológica de los ecosistemas nativos afectando a diversas especies de plantas y animales. Asimismo, la mayoría de las especies depredadoras especialistas son sensibles a la fragmentación o el reemplazo total de los bosques nativos por sistemas agrícolas (Bohac, 1999; Navarrete-Heredia et al., 2002; Klimaszewski et al., 2018). Por otro lado, la remoción de hojarasca y la exposición al suelo desnudo afectan a los escarabajos vagabundos debido, probablemente, a la reducción de la humedad del suelo, lo que los hace propensos a la desecación (Klimaszewski et al., 2018). Por su sensibilidad al cambio de hábitat y a las condiciones microclimáticas en el suelo (Bohac, 1999; Klimaszewski et al., 2018; Méndez-Rojas et al., 2021), estos estafilínidos son considerados indicadores de la perturbación antrópica.
En conclusión, la transformación de los hábitats boscosos por cultivos y pastos reduce la riqueza de especies de escarabajos en diferentes regiones del planeta, donde sólo las especies más generalistas prevalecen en los agroecosistemas (Méndez-Rojas et al., 2021). La expansión urbana también afecta a estos escarabajos, en particular a las especies con hábitos saprófagos que dependen de la alta humedad de los bosques para vivir (Magura et al., 2013). Las razones anteriores hacen importante visibilizar y divulgar la existencia de grupos como los estafilínidos, ya que sólo podemos conservar lo que realmente conocemos, y a pesar de su pequeño tamaño y lo poco llamativo de su coloración, los escarabajos son de gran importancia para el mantenimiento y la funcionalidad de los ecosistemas.
Referencias
- Bohac, J. (1999). Staphylinid beetles as bioindicators. Agriculture, Ecosystems Environment, 74(1-3), 357-372.https://doi.org/10.1016/S0167-8809(99)00043-2.
- Frank, J. H., y Thomas, M. C. (2002). Rove beetles of Florida, Staphylinidae (Insecta: Coleoptera: Staphylinidae). EDIS, (8), 1-13.
- Good, J., y Giller, P. S. (1991). The diet of predatory Staphylinid beetles – a review of records. Entomologist’s Monthly Magazine, 127, 77-89.
- Halimov, F. (2020). Seasonal dynamics of dominant species of soil predators (Coleoptera: Carabidae, Staphylinidae) in agrolandscapes and their potential gluttony. Travaux du Muséum National d’Histoire Naturelle “Grigore Antipa”, 63 (2), 175-187.
- Irmler, U., y Lipkow, E. (2018). Chapter 7. Effect of environmental conditions on distribution patterns of rove beetles. En Betz, O., Irmler, U., y Klimaszewski, J (eds), Biology of Rove Beetles (Staphylinidae) (pp. 117-144). Cham: Springer International Publishing.
- Klimaszewski, J., Brunke, A. J., Work, T. T., y Venier, L. (2018). Chapter 9. Rove beetles (Coleoptera, Staphylinidae) as bioindicators of change in boreal forests and their biological control services in agroecosystems: Canadian case studies. En Betz, O., Irmler, U., y Klimaszewski, J (eds), Biology of Rove Beetles (Staphylinidae) (pp. 161-181). Cham: Springer International Publishing.
- López-García, M. M., y Marín-Gómez, O. H. (2018). Description and notes on natural history of a new species of Parosus Sharp, 1887 (Coleoptera, Staphylinidae, Oxytelinae) living in floral bracts of Columnea medicinalis L. (Gesneriaceae). Zootaxa, 4394 (4), 559-566. https://doi.org/10.11646/zootaxa.4394.4.6.
- Magura, T., Nagy, D., y Tóthmérész, B. (2013). Rove beetles respond heterogeneously to urbanization. Journal of Insect Conservation, 17(4), 715-724. https://doi.org/10.1007/s10841-013-9555-y.
- Márquez, J., y Navarrete-Heredia, J. L. (1994). Especies de Staphylinidae (Insecta, Coleoptera) asociadas a detritos de Atta mexicana (Hymenoptera, Formicidae) en dos localidades de Morelos, México. Folia Entomológica Mexicana, 91, 31-46.
- Méndez-Rojas, D. M., Cultid-Medina, C., y Escobar, F. (2021). Influence of land use change on rove beetle diversity: A systematic review and global meta-analysis of a mega-diverse insect group. Ecological Indicators, 122, 107239.
- Moreira, I. E., Pires-Silva, C. M., Ribeiro, K. G., Zilberman, B., y Bezerra-Gusmão, M. A. (2019). Run to the nest: A parody on the Iron Maiden song by Corotoca spp. (Coleoptera, Staphylinidae). Papéis Avulsos de Zoologia, 59, 1-6. https://doi.org/10.11606/1807-0205/2019.59.18.
- Navarrete-Heredia, J. L., Newton, A. F., Thayer, M. K., Ashe, J. S., y Chandler, D. S. (2002). Guía ilustrada para los géneros de Staphylinidae (Coleoptera) de México (Issue C/595.760972 G8). Universidad de Guadalajara, CONABIO.
- Parker, J. (2017). Staphylinids. Current Biology, 27 (2), R49-R51.
- Pohl, G. R., Langor, D. W., y Spence, J. R. (2007). Rove beetles and ground beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of harvest and regeneration practices in western Canadian foothills forests. Biological Conservation, 137(2), 294-307.
- Sayers, T. D., Steinbauer, M. J., y Miller, R. E. (2019). Visitor or vector? The extent of rove beetle (Coleoptera: Staphylinidae) pollination and floral interactions. Arthropod-Plant Interactions, 13 (5), 685-701. https://doi.org/10.1007/s11829-019-09698-9.
- Silva, F. S., Lima, D. C. B., Costa, A. A. V., y Campos, D. S. L. (2014). An ethno-entomological study of rove beetles of the genus Paederus (Coleoptera, Staphylinidae) in the municipality of Chapadinha, Maranhao state, notheastern Brazil. Boletín de la Sociedad de Entomología Aragonesa, (55), 351-355.
- Von Beeren, C., Brückner, A., Maruyama, M., Burke, G., Wieschollek, J., y Kronauer, D. J. (2018). Chemical and behavioral integration of army ant-associated rove beetles–a comparison between specialists and generalists. Frontiers in Zoology, 15 (1), 1-16. https://doi.org/10.1186/s12983-018-0249-x.
Recepción: 08/09/2021. Aprobación: 09/12/2022.